Tema 2.- Cormofitas: del Silúrico al
Carbonífero
| Carlos Romero Zarco, Universidad de Sevilla |
Características
- Origen - Ciclo
isospórico - Devónico
Inferior - Otros fósiles del Silúrico al
Devónico - Del Devónico al Carbonífero -
Lycopodiopsida
- Polypodiopsida
- Progimnospermas
- Sistemas de
reproducción sexual en los helechos del Carbonífero - Ciclo
heterospórico - Resumen
Índice de
temas - Tema anterior
- Tema siguiente -
| Origen de las plantas terrestres |
|
|
|
|
|
|
|
 Hipótesis de Jefrey (de Scagel) |
|
|
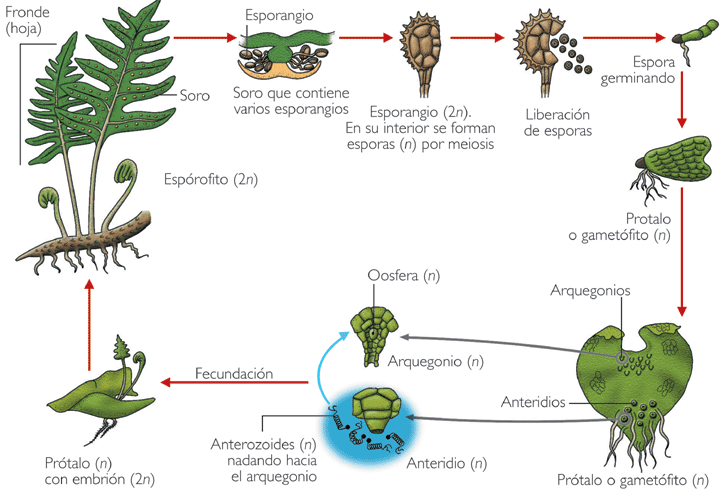 Ciclo de vida de un helecho isospórico actual |
| El registro fósil
del Devónico
inferior (410 m.a.): Riniales (Rhyniales) |
|
|
|
|
|
|
|
 Aglaophyton major
|
 Rhynia gwynne-vaughanii
|
| De
Falconaumanni - Trabajo propio, CC BY-SA 3.0,
https://commons.wikimedia.org/w/index.php?curid=36961871 |
De
Falconaumanni - Trabajo propio, CC BY-SA 3.0,
https://commons.wikimedia.org/w/index.php?curid=36961871 |
 Flora fósil del yacimiento Rhynie Chert (esquitos, Escocia, hace 410 m.a.). A. Rhynia gwynne-vaughanii. B.
Aglaophyton major. C. Ventarura lyonii.
D. Asteroxylon mackiei. E. Horneophyton
lignieri. F. Nothia aphylla.
De
Falconaumanni - Trabajo propio, CC BY-SA 3.0,
https://commons.wikimedia.org/w/index.php?curid=37260627 |
|
|
|
|
|
|
|
 |
Corte transversal de un tallo de Rhynia mostrando la pequeña protostela central. De Plantsurfer - Trabajo propio, CC BY-SA 2.0 uk, https://commons.wikimedia.org/w/index.php?curid=3823922 Al parecer, el xilema de las Riniales era de maduración centrífuga (vasos maduros periféricos, al igual que las plantas actuales). Todas las riniales se extinguieron durante el Devónico. |
| Otros fósiles del Silúrico y
del Devónico |
|
 Zosterophyllum (Zosterofilales),
final del Silúrico-Devónico Inferior. Xilema centrípeto.
|
 Psilophyton (Trimerofitales),
Devónivo Medio. Xilema centrífugo.
|
| Se cree que
las Zosterofilales dieron origen a las "Licopodiofitas"
(Cl. Lycopodiopsida) |
Se cree que
las Trimerofitales dieron origen a las Equisetopsida,
las Polypodiopsida y a las Espermatofitas |
| Del Devónico
al Carbonífero |
|
|
|
|
|
|
|
|
|
|
|
 Lepidodendrales del Carbonífero De Falconaumanni - Trabajo propio, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=46906747 |
|
|
|
|
|
|
|
|
|
|
|
 |
 |
| Tronco de Lepidodendron
mostrando las cicatrices foliares. By Verisimilus T - Own work (Original text: self-made), CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=6741052 |
Fragmento de Sigillaria.
De Jstuby de Wikipedia en inglés - Trabajo propio, Dominio público, https://commons.wikimedia.org/w/index.php?curid=17781090 |
 |
A la izquierda tronco y raíces principales de Sigillaria, el famoso Stanhope-tree (c. -320 m.a.), en el N de Inglaterra. El árbol pudo tener unos 30 m de altura. De Ashley Dace, CC BY-SA 2.0, https://commons.wikimedia.org/w/index.php?curid=16062364 |
|
|
 |
 Tronco fósil de Calamithes (Villanueva del Río y Minas). Foto de la colección de Jaime Gil Fernández https://www.cepalcala.org/ciencias1/geologia/paleontologia/paleobotanica/fosiles_vegetales_sevilla_1.htm A la izquierda, reconstrucción de planta y hojas de Calamites. By Falconaumanni - Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=56797814 |
|
|
|
|
|
|
|
|
|
|
|
|
 Por Jeanne Morgan - https://commons.wikimedia.org |
 Hojas de tipo "Pecopteris" del
Carbonífero.
By Woudloper - Own work,
Public Domain,
https://commons.wikimedia.org/w/index.php?curid=3370795
|
|
|
  |
 |
|
|
|
|
| Sistemas de reproducción sexual de
los helechos del Carbonífero |
|
|
|
|
|
|
|
|
|
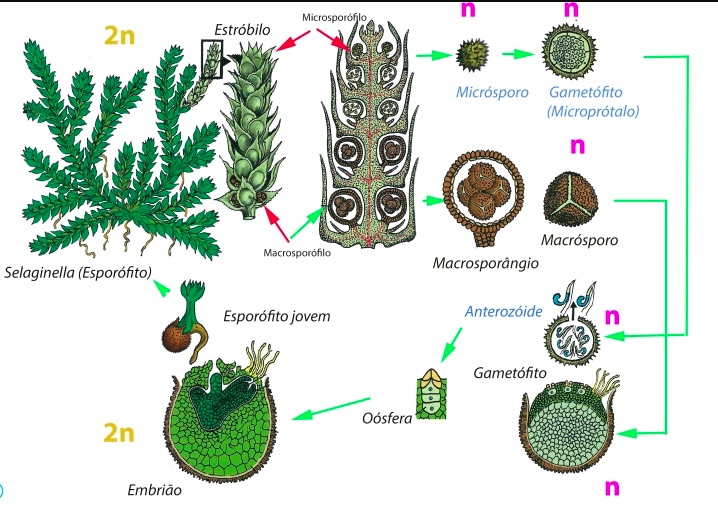 Ciclo de vida de un helecho heterospórico actual: Selaginella |
|
| RESUMEN |
|



{kind=link}